
Embryological Development and Anatomy of the Human Heart
1. Toichieva Zarina Zhamaldinovna
2. Altamas Khan
Sujal Soni
(1. Lecturer, International Medical Faculty, Osh State University, Osh, Kyrgyz Republic
2. Students, International Medical Faculty, Osh State University, Osh, Kyrgyz Republic)
Abstract
The human heart is the first organ to become functional in the developing embryo, beginning to beat as early as the third week of gestation, long before most other organ systems have even begun to take shape. This early functionality is not a coincidence — the embryo quickly outgrows simple diffusion as a means of nutrient and oxygen delivery, and a working circulation becomes a survival necessity within days. This narrative review walks through cardiac embryology in the order it actually happens cardiogenic mesoderm formation, primitive heart tube formation, looping, septation, and outflow tract remodelling and at each stage links the developmental event to the adult anatomy it produces, and to the congenital defect that results when that particular step fails. The aim is to fill a gap that most textbooks leave unaddressed: a single continuous narrative connecting embryology directly to adult cardiac anatomy and to clinically relevant congenital heart disease, rather than treating the two as separate chapters.
Keywords: cardiogenic mesoderm, heart tube, cardiac looping, septation, endocardial cushions, congenital heart disease
1. Introduction
Most students first meet cardiac embryology as a confusing sequence of folds, loops, and septa, memorised only long enough to clear an exam and forgotten soon after. But cardiac embryology is, in fact, one of the best-organised stories in all of developmental anatomy, because every single congenital heart defect a clinician encounters later in life is simply a freeze-frame of a normal developmental step that did not complete properly. A ventricular septal defect is not a random hole — it is the membranous part of the interventricular septum that failed to close on schedule. Transposition of the great arteries is not a random vessel mix-up — it is a failure of the normal spiralling of the aorticopulmonary septum. Once a student sees embryology this way, the whole subject becomes far easier to retain, because anatomy and pathology start explaining each other.
This review tries to present cardiac development in exactly that spirit — as a single continuous timeline from the third week of gestation to the fully septated four-chambered heart, pausing at each stage to connect the embryological event with the adult structure it leaves behind, and with the defect that results if the step is incomplete.
2. Formation of the Cardiogenic Field
2.1 The First and Second Heart Fields
Cardiac development begins around day 15–16 of gestation, when a horseshoe-shaped region of splanchnic mesoderm called the cardiogenic field (or cardiogenic crescent) becomes specified, lying cranial to the prochordal plate and the buccopharyngeal membrane. This cardiogenic field is not a single uniform population of cells — it is now understood to be composed of two distinct but overlapping progenitor populations:
The First Heart Field (FHF) — gives rise mainly to the left ventricle and parts of both atria. These are the earliest cardiac progenitors to differentiate.
The Second Heart Field (SHF) — a population lying medial and dorsal to the FHF, which contributes later and continues adding cells to the elongating heart tube, particularly forming the right ventricle and the outflow tract.
This two-field model is important clinically because most outflow tract congenital anomalies (such as tetralogy of Fallot and transposition of the great arteries) trace back to disturbances of the SHF and its associated neural crest contribution, rather than to the FHF.
2.2 Lateral Folding and Tube Formation
As the embryo undergoes lateral body folding, the originally flat, horseshoe-shaped cardiogenic crescent is pulled ventrally and brought together in the midline, fusing the paired endocardial heart tubes into a single primitive heart tube by around day 21–22. This tube is suspended within the pericardial cavity by a dorsal mesocardium, most of which later breaks down except at the arterial and venous poles, allowing the tube freedom to loop in the following stage.
The wall of this early heart tube already shows the three layers that persist into adult cardiac histology:
Endocardium — derived from the endothelial-lined inner tube, becomes the adult endocardium.
Myocardium — the surrounding muscular mantle, becomes the adult myocardium.
Epimyocardium/visceral pericardium — later contributed to substantially by cells from the proepicardial organ, becomes the adult epicardium.
Between the endocardium and myocardium lies a gelatinous extracellular matrix called cardiac jelly, which is critical later on because it provides the substrate into which cells migrate to form the endocardial cushions.
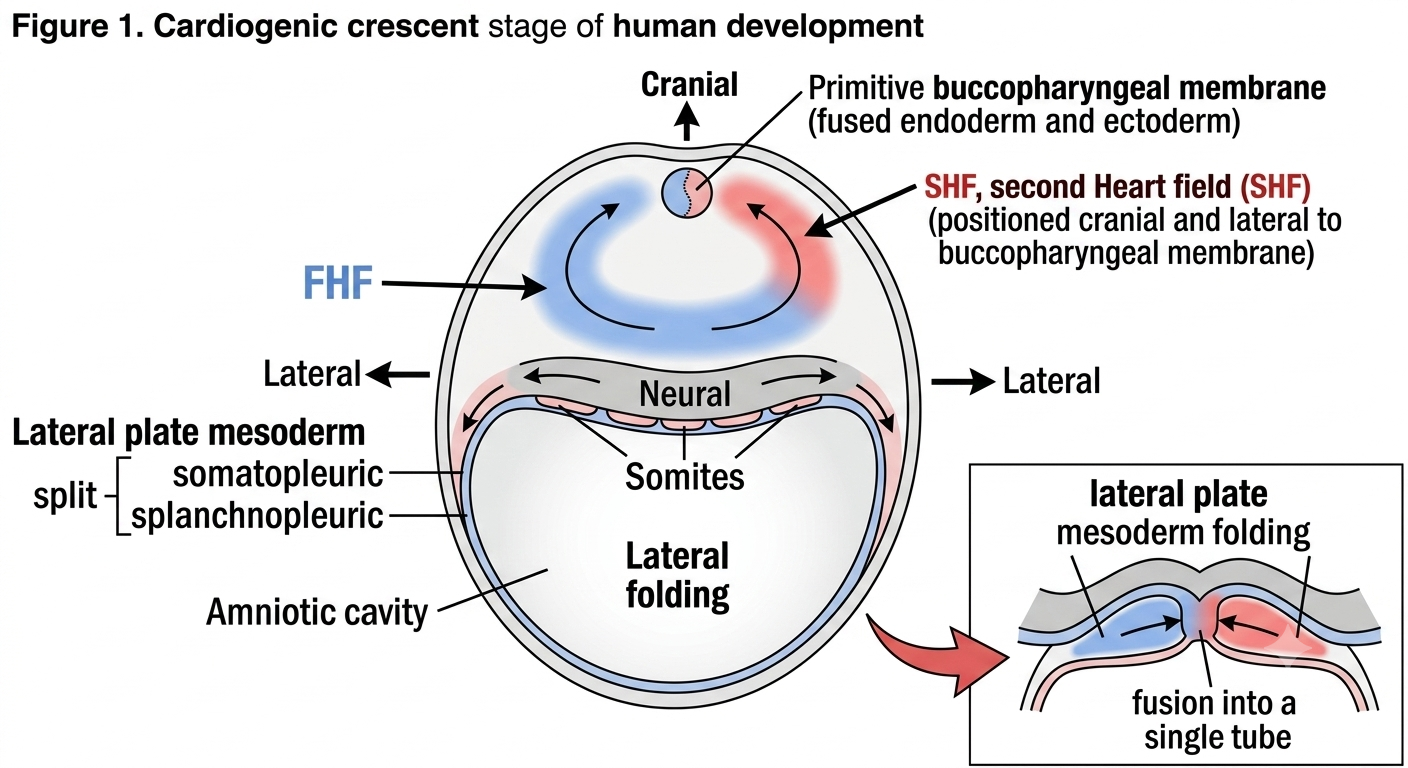
Figure 1. A dorsal view of the embryological disc at approximately day 18, showing the crescent-shaped cardiogenic region located cranial and lateral to the buccopharyngeal membrane.
3. The Primitive Heart Tube: Five Segments
By around day 23, the straight primitive heart tube has differentiated, from cranial (arterial) to caudal (venous) end, into five recognisable dilatations:
Truncus arteriosus — will become the proximal aorta and pulmonary trunk after spiral septation.
Bulbus cordis — divided into a conus (which forms the outflow portions/infundibula of both ventricles) and a proximal part contributing to the right ventricle.
Primitive ventricle — forms the bulk of the definitive left ventricle.
Primitive atrium — forms the trabeculated parts of both definitive atria.
Sinus venosus — receives the vitelline, umbilical, and common cardinal veins; its right horn is incorporated into the right atrium and gives rise to the sinoatrial node region; its left horn regresses to become the coronary sinus.
This cranio-caudal naming convention is genuinely useful to memorise, because every adult structure can be traced back to exactly one of these five segments, and most textbook diagrams of "heart development" are simply showing this same tube being bent, twisted, and partitioned.
Clinical Correlation: The Sinoatrial Node
Because the sinoatrial node develops from the right horn of the sinus venosus as it is incorporated into the right atrial wall, the natural cardiac pacemaker ends up sitting near the junction of the superior vena cava and right atrium in the adult — exactly where it is found on dissection and on ECG-guided lead placement. This single embryological fact explains the entire adult anatomical position of the conduction system's primary pacemaker.
4. Cardiac Looping
4.1 The Process
Around day 23–28, the straight heart tube begins to loop. Because both ends of the tube (the arterial and venous poles) are relatively fixed in position while the tube itself continues to elongate rapidly, the only way for the extra length to be accommodated is by bending. The bulboventricular portion of the tube bends ventrally, caudally, and to the right, while the atrial and sinus venosus portion shifts dorsally, cranially, and to the left. This establishes the first visible left-right asymmetry in the human body and determines the normal dextral (rightward) looping pattern, abbreviated as D-looping.
4.2 Why Looping Matters
Looping is not a cosmetic step — it is what brings the future right ventricle (from the bulbus cordis) to lie anteriorly and to the right of the future left ventricle (from the primitive ventricle), exactly as it sits in the adult chest. Without looping, the heart would remain a straight tube incapable of generating the spatial arrangement needed for the chambers to later connect correctly to their respective great vessels.
Clinical Correlation: Situs and Looping Defects
When looping occurs in the mirror-image direction (L-looping) instead of the normal D-looping, the result is dextrocardia or related disorders of cardiac situ. These conditions are frequently, though not always, associated with more complex congenital heart disease, because looping direction also influences how the atrioventricular and ventriculo-arterial connections subsequently align.
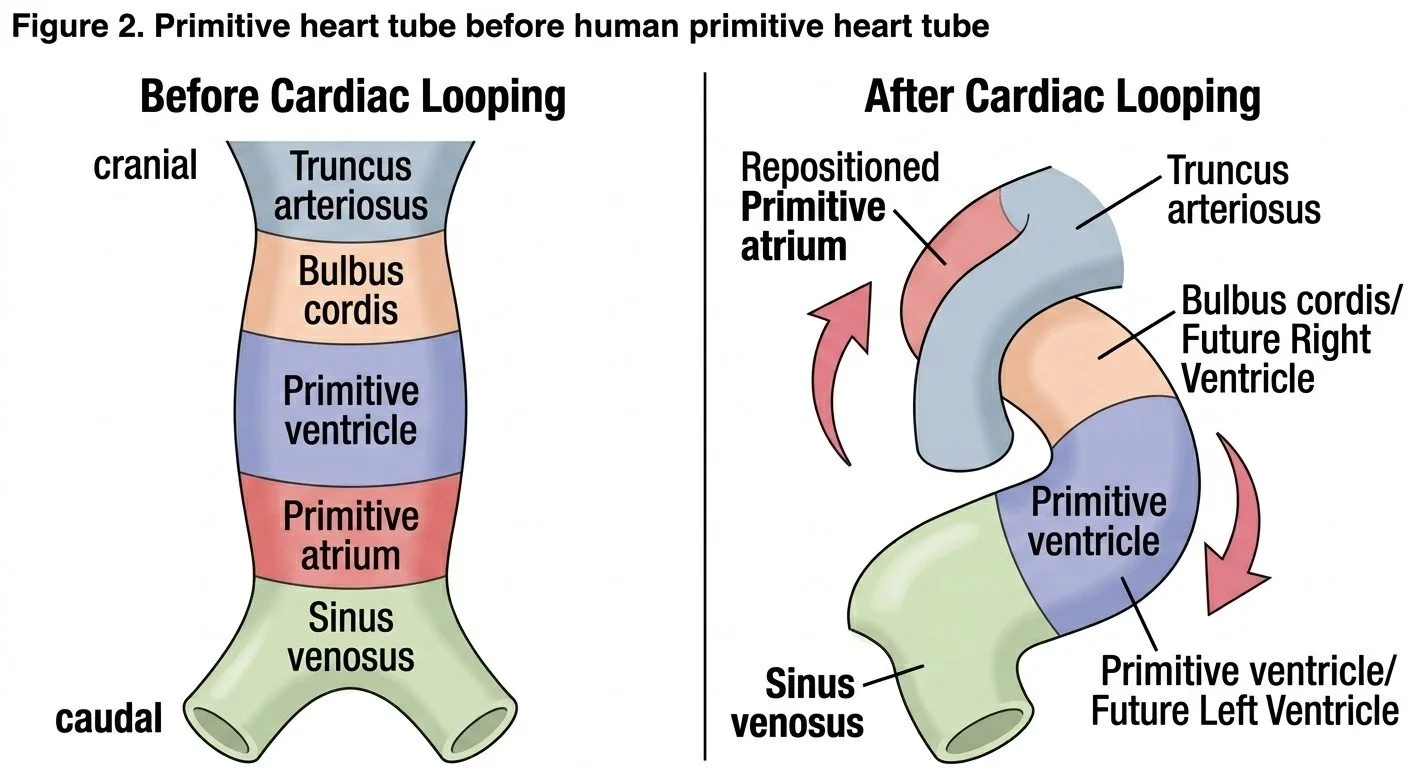
Figure 2. An intermediate stage, showing lateral folding and the fusion of the paired heart-forming regions into a single, midline, linear heart tube, with its segments color-coded to indicate their eventual adult fates (truncus arteriosus, bulbus cordis, primitive ventricle, primitive atrium, sinus venosus).
5. Formation of the Cardiac Chambers
As looping completes, the originally simple segments begin to "balloon" outward at specific points to form the definitive chambers, while the original primitive segments are retained internally as the smooth-walled "inlet" and trabeculated "outlet" portions that anatomists still identify in the adult heart. This is why the adult right atrium has a smooth posterior wall (derived from the sinus venosus, the sinus venarum) and a rough anterior trabeculated wall (derived from the primitive atrium) — two different embryological origins fused into a single adult chamber, separated internally by the crista terminalis. The same dual origin explains the smooth versus trabeculated portions of both ventricles.
By approximately day 28, all four chambers are recognisable in their correct relative positions, even though septation is still incomplete.
6. Atrial Septation
Atrial septation is one of the most clinically important stages, because it must accomplish something tricky — dividing the atrium into two chambers while still keeping a right-to-left shunt open, since fetal circulation depends on bypassing the non-functional fetal lungs. This is achieved through a sequence of three septa/openings, often summarised as the "septum primum, septum secundum" story:
The septum primum grows down from the roof of the common atrium toward the fusing endocardial cushions, leaving a gap below it called the ostium primum.
Before the ostium primum closes completely (as the septum primum fuses with the cushions), a new opening called the ostium secundum appears higher up within the septum primum itself, maintaining the shunt.
A second, thicker septum, the septum secundum, grows down to the right of the septum primum, but it too has an opening — the foramen ovale — which is positioned so that the flap-like septum primum overlies it like a one-way valve, allowing blood to pass right-to-left but not left-to-right.
At birth, when pulmonary vascular resistance drops and left atrial pressure rises above right atrial pressure, the septum primum is pressed against the septum secundum, functionally closing the foramen ovale; anatomical fusion follows over the following months to a year, leaving the adult fossa ovalis as the permanent scar of this entire process.
Clinical Correlation: Atrial Septal Defects
An ostium secundum ASD, the most common type, results from excessive resorption of the septum primum or an undersized septum secundum, leaving a persistent communication even after birth. An ostium primum ASD, by contrast, results from failure of the septum primum to fuse with the endocardial cushions, and is therefore frequently associated with atrioventricular valve abnormalities, because the same cushion tissue forms both structures. Recognising which embryological step failed immediately tells a student which associated anomalies to expect on echocardiography.
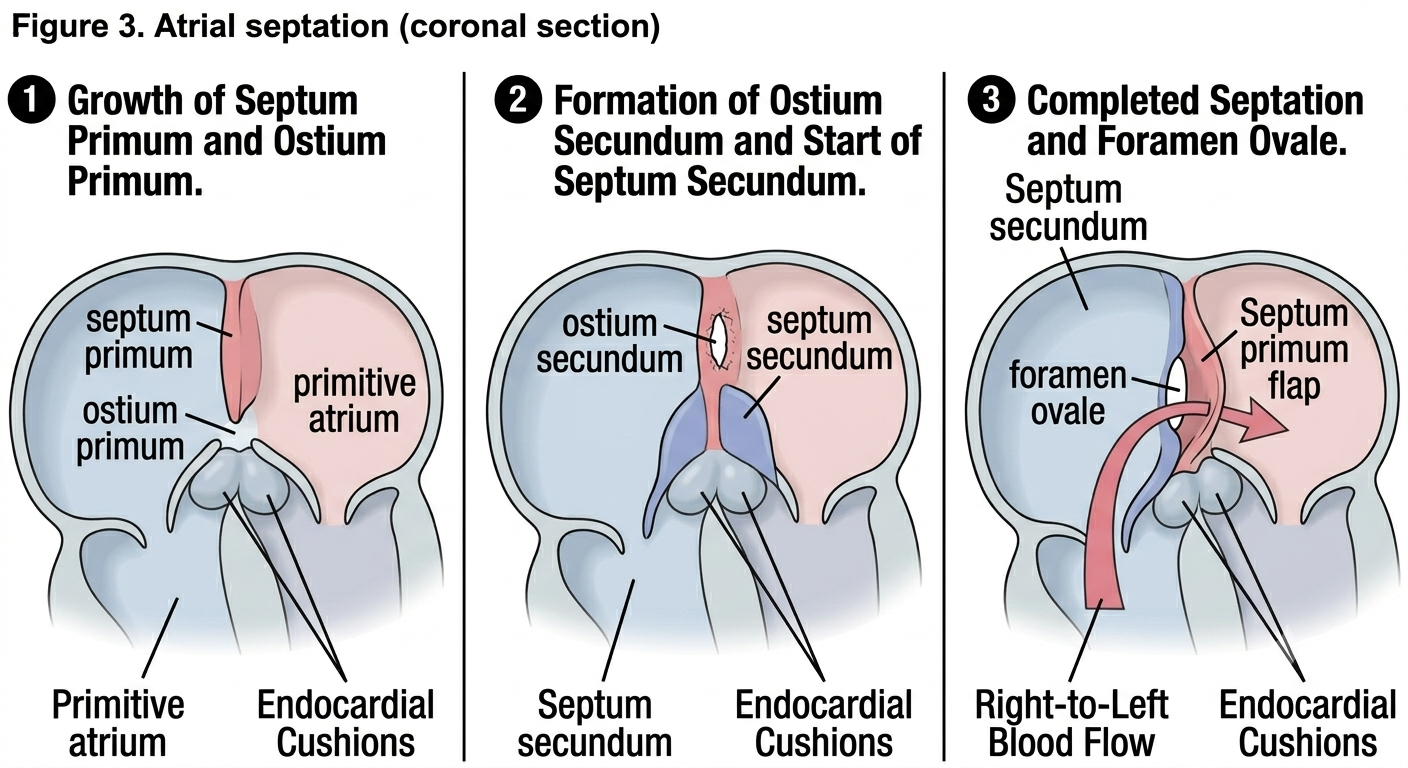
Figure 3. A three-panel illustration detailing the complex process of atrial partitioning. (1) Growth of the septum primum and formation of the ostium primum. (2) Breakdown of the septum primum (ostium secundum) and subsequent growth of the septum secundum. (3) Overlapping of the septa to form the foramen ovale, with the septum primum acting as a valve allowing right-to-left blood flow.
7. The Endocardial Cushions
The endocardial cushions are localised swellings of cardiac jelly, subsequently invaded by mesenchymal cells (derived in part from an epithelial-to-mesenchymal transition of the overlying endocardium), found at two key sites — the atrioventricular canal and the outflow tract. Their importance can hardly be overstated, because they contribute to almost every septal and valvular structure in the mature heart:
Fusion of the superior and inferior atrioventricular cushions divides the single atrioventricular canal into separate right and left atrioventricular orifices, and contributes to the membranous part of the interventricular septum and the lower part of the atrial septum.
The cushions also give rise to the leaflets of the mitral and tricuspid valves, in conjunction with surrounding myocardial tissue that later undergoes delamination.
A separate, spiralling set of outflow tract cushions divides the truncus arteriosus and bulbus cordis into the aorta and pulmonary trunk, discussed further below.
Clinical Correlation: Atrioventricular Septal Defects
Failure of the superior and inferior atrioventricular cushions to fuse produces an atrioventricular septal defect (previously called endocardial cushion defect), in which there is a common atrioventricular valve and a combined atrial and ventricular septal defect. This anomaly is strongly associated with Down syndrome, illustrating how a single embryological structure, when disturbed, produces a recognisable and predictable cluster of anatomical defects rather than a random assortment.
8. Ventricular Septation
The interventricular septum forms through the combined growth of two distinct components, and recognising this dual origin is essential to understanding why some ventricular septal defects are muscular and others are membranous:
A muscular interventricular septum grows upward from the floor of the common ventricle, formed largely by myocardium from both the primitive ventricle and bulbus cordis as they expand and meet in the midline.
A membranous interventricular septum is a much smaller, thinner component formed from tissue derived from the fused endocardial cushions (along with contributions from the outflow tract cushions), closing the residual interventricular foramen that remains after the muscular septum has grown most of the way up.
Clinical Correlation: Ventricular Septal Defects
Ventricular septal defects are the most common congenital heart defects overall, and the overwhelming majority are membranous, precisely because the membranous septum is the smallest component, formed last, and dependent on the successful fusion of multiple separate tissue contributions (endocardial cushion tissue plus outflow cushion tissue) — more steps mean more opportunity for something to go slightly wrong. Muscular VSDs are less common and tend to be multiple ("Swiss cheese septum") because the muscular septum forms by the coalescence of several separate trabeculated outgrowths rather than a single sheet.
9. Outflow Tract Septation and the Role of Neural Crest
9.1 Spiral Septation
The truncus arteriosus and bulbus cordis are divided into the aorta and pulmonary trunk by a pair of outflow tract (truncoconal) cushions that grow toward each other and fuse along a spiral course, rather than a straight line. This spiral fusion is the reason the aorta and pulmonary trunk wind around one another in the adult chest rather than running as two parallel straight tubes — the spiral path of septation is directly responsible for the spiral anatomical relationship of the two great vessels at the base of the heart.
9.2 Cardiac Neural Crest Contribution
Crucially, this spiral outflow septation is not accomplished by cardiac mesoderm alone. A specialised population of cardiac neural crest cells, originating from the dorsal neural tube at the level of the developing hindbrain, migrates through the pharyngeal arches and invades the outflow tract cushions, where it is essential for proper formation of the aorticopulmonary septum, the semilunar valves, and the smooth muscle of the proximal great vessels. Disruption of this neural crest population — whether by genetic mutation, teratogen exposure, or a primary migration defect — produces a recognisable and clinically important family of outflow tract anomalies.
Clinical Correlation: Conotruncal Anomalies
Persistent truncus arteriosus — complete failure of aorticopulmonary septation, leaving a single common outflow vessel.
Transposition of the great arteries — the spiral septation fails to follow its normal helical course, resulting in the aorta arising from the right ventricle and the pulmonary trunk from the left ventricle.
Tetralogy of Fallot — results from anterosuperior deviation of the outflow septum, producing a malaligned ventricular septal defect, pulmonary outflow obstruction, an overriding aorta, and consequent right ventricular hypertrophy.
Many of these conotruncal anomalies cluster together with craniofacial and thymic/parathyroid abnormalities in syndromes such as DiGeorge syndrome (22q11 deletion), which makes sense once it is appreciated that the same cardiac neural crest population also contributes to pharyngeal arch derivatives such as the thymus and parathyroid glands — one disrupted cell population, multiple seemingly unrelated organ systems affected.
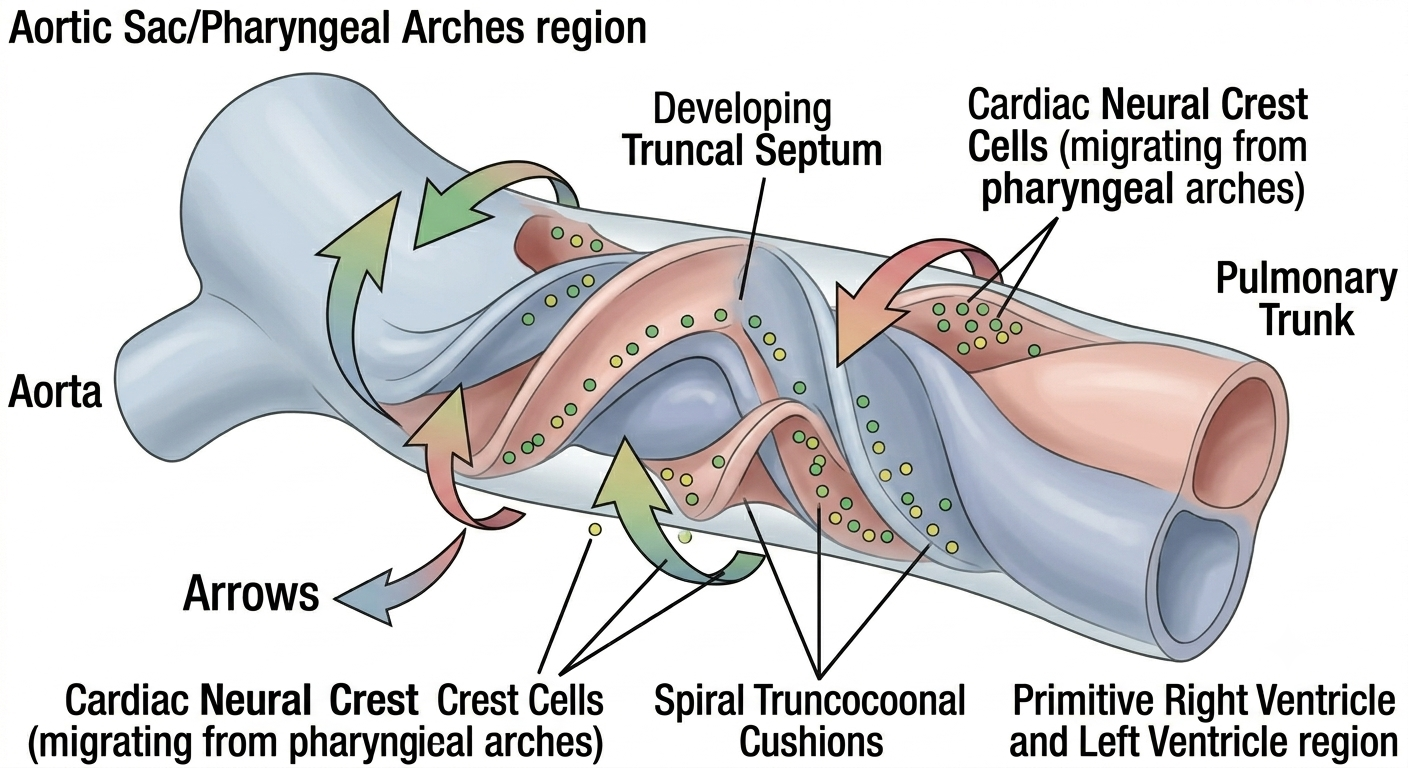
Figure 4. A three-dimensional representation of the developing pharyngeal arch region and the embryonic heart, with color-coded dots illustrating the critical migration of cardiac neural crest cells from the dorsal neural tube to the heart's outflow tract
10. Cardiac Valve Formation
The atrioventricular valves (mitral and tricuspid) form from a combination of endocardial cushion tissue and adjacent ventricular myocardium, which undergoes a process of delamination — the inner layer of ventricular wall is gradually undermined and thinned to form valve leaflets and chordae tendineae, while the outer layer remains as the muscular ventricular wall. The semilunar valves (aortic and pulmonary) form from local swellings within the outflow tract cushions themselves, remodelled into the characteristic three-cusped arrangement seen in the adult.
Clinical Correlation: Bicuspid Aortic Valve
A bicuspid aortic valve, one of the most common congenital cardiac anomalies in adults (often discovered incidentally), results from abnormal fusion of two of the three normal cusp-forming swellings during this remodelling process. Although frequently asymptomatic for decades, it predisposes to later valve calcification, stenosis, and aortic dilation, illustrating how a subtle embryological variation can have a delayed clinical consequence that only becomes apparent well into adult life.
11. Development of the Cardiac Conduction System
Although the conduction system is often taught separately from "structural" cardiac embryology, it develops in close anatomical steps with the chambers themselves. The sinoatrial node, as already mentioned, derives from the right horn of the sinus venosus. The atrioventricular node and the bundle of His are thought to arise from a ring of specialised, slow-conducting myocardium around the atrioventricular canal — essentially leftover primitive "nodal-type" myocardium that fails to fully transform into fast-conducting working myocardium, retaining instead the slow, automatic electrical properties characteristic of the original primitive heart tube before the more rapidly conducting chamber myocardium differentiated around it.
This developmental detail nicely explains a basic electrophysiological fact every student is taught — the AV node normally conducts slowly compared to the rest of the heart, which from this embryological perspective is simply primitive-type tissue that never "upgraded."
12. Fetal Circulation and the Transition at Birth
No discussion of cardiac embryology is complete without fetal circulation, because the heart's job before birth is fundamentally different from its job after birth — before birth, the lungs do not oxygenate blood, the placenta does, and the entire circulatory plan is built around three key shunts that bypass the non-functional fetal lungs and the immature fetal liver:
Ductus venosus — shunts oxygenated blood from the umbilical vein largely past the liver into the inferior vena cava.
Foramen ovale — shunts blood from the right atrium directly into the left atrium, bypassing the right ventricle and lungs (as described in Section 6).
Ductus arteriosus — shunts blood from the pulmonary trunk directly into the descending aorta, bypassing the lungs.
At birth, clamping of the umbilical cord and the first breaths trigger a cascade of pressure changes — pulmonary vascular resistance falls sharply, systemic vascular resistance rises, left atrial pressure exceeds right atrial pressure, and prostaglandin levels fall (removing the chemical signal that had kept the ductus arteriosus open). All three shunts close, functionally within minutes to hours and anatomically over subsequent weeks to months, completing the transition to the adult circulatory pattern.
Clinical Correlation: Patent Ductus Arteriosus
When the ductus arteriosus fails to close, usually because of persistently low oxygen tension or abnormal prostaglandin signalling (more common in premature infants), the result is a patent ductus arteriosus, producing a continuous "machinery" murmur and a left-to-right shunt once pulmonary resistance has fallen below systemic resistance. Indomethacin or ibuprofen, both prostaglandin synthesis inhibitors, are used pharmacologically to encourage closure — a direct clinical exploitation of the same prostaglandin pathway that keeps the duct open in utero.
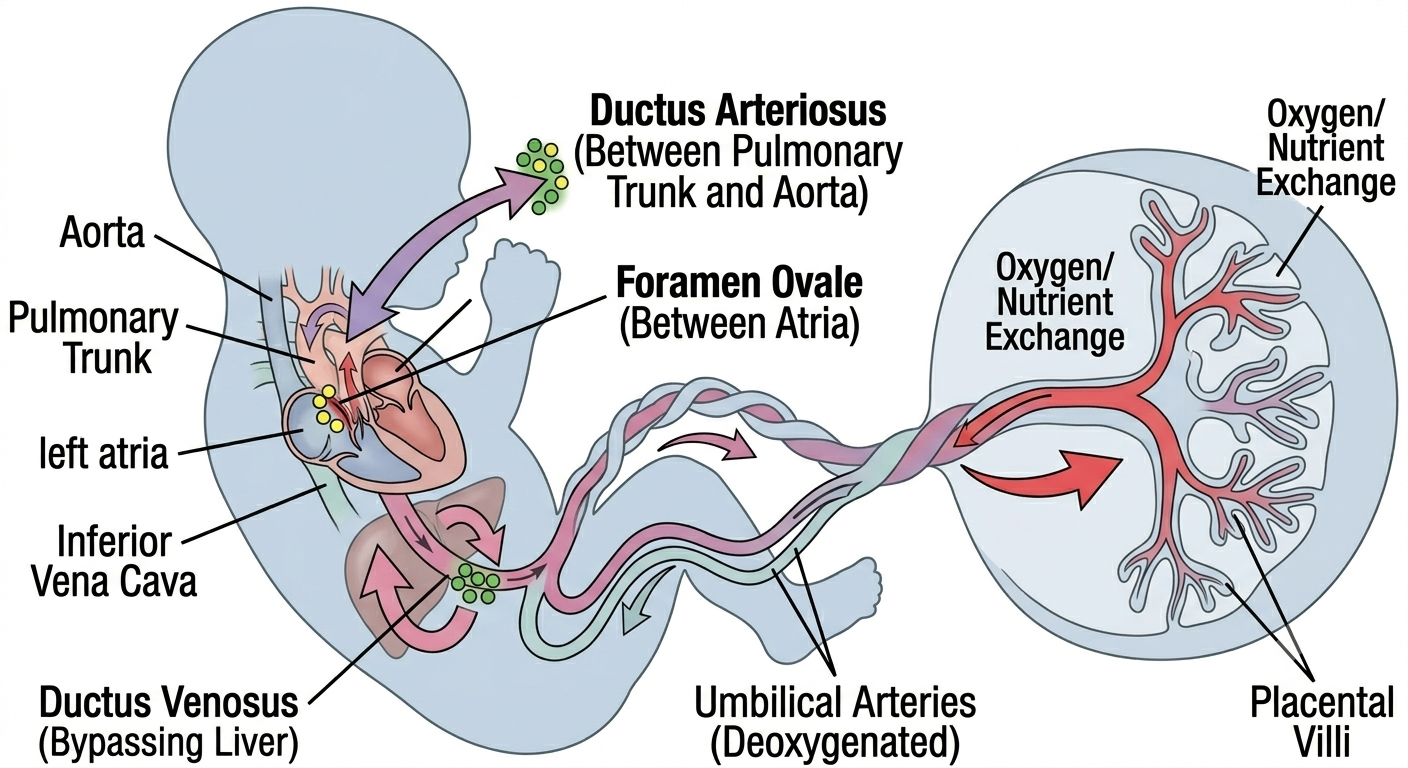
Figure 5. A summary infographic showing the unique characteristics of the fetal circulatory system, including the placenta, the composition of umbilical vessels, and the major physiological shunts (ductus venosus, foramen ovale, ductus arteriosus) that adapt to parallel pumping before the lungs are operational.
13. From Embryology to Adult Topography: A Summary Map
It is worth closing with a short summary connecting embryological origin directly to adult anatomical landmark, since this is ultimately the most useful revision tool this review can offer:
Right atrium smooth wall (sinus venarum) → sinus venosus.
Right atrium trabeculated wall and pectinate muscles → primitive atrium.
Crista terminalis → boundary between the above two embryological territories.
Fossa ovalis → remnant of the septum primum within the closed foramen ovale.
Smooth-walled inflow of right ventricle → bulbus cordis (proximal part).
Conus/infundibulum of right ventricle → bulbus cordis (conal part).
Trabeculated left ventricle → primitive ventricle.
Membranous interventricular septum → fused endocardial cushion tissue.
Coronary sinus → left horn of sinus venosus.
Ligamentum arteriosum (adult remnant) → closed ductus arteriosus.
Ligamentum teres of the liver and ligamentum venosum → remnants of the umbilical vein and ductus venosus respectively.
14. Conclusion
Cardiac embryology is sometimes treated as a separate, almost decorative chapter in anatomy courses, disconnected from the "real" adult anatomy that students are eventually tested on clinically. This review has tried to argue, through each developmental stage, that this separation is artificial. Every adult landmark in the heart — the crista terminalis, the fossa ovalis, the membranous septum, the spiral relationship of the great vessels — is a direct embryological signature, and every major congenital heart defect is simply that same developmental step caught incomplete. Approaching cardiac anatomy through its embryological history, rather than as a static finished structure, gives a student a far more durable and clinically useful understanding than memorisation of either subject in isolation.
References
1. Hikspoors JPJM, Lamers WH, Kerwin J, Hu Z, Henderson DJ, Anderson RH. Relating normal human cardiac development to the anatomical findings in the congenitally malformed heart. Clin Anat. 2025 Apr;38(3):296-313. doi: 10.1002/ca.24240. Epub 2024 Nov 13.
2. Buijtendijk MFJ, Barnett P, van den Hoff MJB. Development of the human heart. Am J Med Genet C Semin Med Genet. 2020 Mar;184(1):7-22. doi: 10.1002/ajmg.c.31778. Epub 2020 Feb 12. PMID: 32048790; PMCID: PMC7078965.
3. Sylva M, van den Hoff MJB, Moorman AFM. Development of the human heart. Am J Med Genet A. 2014;164A(6):1347-1371.
4. Moorman AFM, Christoffels VM. Cardiac chamber formation: development, genes, and evolution. Physiol Rev. 2003;83(4):1223-1267.
5. Männer J. Cardiac looping in the chick embryo: a morphological review with special reference to terminological and biomechanical aspects of the looping process. Anat Rec. 2000;259(3):248-262.
6. Srivastava D, Olson EN. A genetic blueprint for cardiac development. Nature. 2000;407(6801):221-226.
7. Kirby ML, Gale TF, Stewart DE. Neural crest cells contribute to normal aorticopulmonary septation. Science. 1983;220(4601):1059-1061.
8. Hutson MR, Kirby ML. Neural crest and cardiovascular development: a 20-year perspective. Birth Defects Res C Embryo Today. 2003;69(1):2-13.
9. Keyte A, Hutson MR. The neural crest in cardiac congenital anomalies. Differentiation. 2012;84(1):25-40.
10. Plein A, Fantin A, Ruhrberg C. Neural crest cells in cardiovascular development. Curr Top Dev Biol. 2015;111:183-200.
11. Bartman T, Hove J. Mechanics and function in heart morphogenesis. Dev Dyn. 2005;233(2):373-381.
12. Taber LA. Mechanical aspects of cardiac development. Prog Biophys Mol Biol. 2006;90(1-3):75-100.
13. Gittenberger-de Groot AC, Bartelings MM, Deruiter MC, Poelmann RE. Basics of cardiac development for the understanding of congenital heart malformations. Pediatr Res. 2005;57(2):169-176.
14. Anderson RH, Brown NA, Moorman AFM. Development and structures of the venous pole of the heart. Dev Dyn. 2006;235(1):2-9.
15. Anderson RH, Webb S, Brown NA, Lamers W, Moorman A. Development of the heart: (2) septation of the atriums and ventricles. Heart. 2003;89(8):949-958.
16. Webb S, Brown NA, Anderson RH. Formation of the atrioventricular septal structures in the normal mouse heart. Circ Res. 1998;82(6):645-656.
17. Snarr BS, Kern CB, Wessels A. Origin and fate of cardiac mesenchyme. Dev Dyn. 2008;237(10):2804-2819.
18. Person AD, Klewer SE, Runyan RB. Cell biology of cardiac cushion development. Int Rev Cytol. 2005;243:287-335.
19. Combs MD, Yutzey KE. Heart valve development: regulatory networks in development and disease. Circ Res. 2009 Aug 28;105(5):408-421. doi: 10.1161/CIRCRESAHA.109.201566. PMID: 19713546; PMCID: PMC2777683.
20. Moreno-Rodriguez RA, Krug EL, Reyes L, Villavicencio L, Mjaatvedt CH, Markwald RR. Bidirectional fusion of the heart-forming fields in the developing chick embryo. Dev Dyn. 2006;235(1):191-202.
21. Lamers WH, Moorman AFM. Cardiac septation: a late contribution of the embryonic primary myocardium to heart morphogenesis. Circ Res. 2002 Jul 26;91(2):93-103. doi: 10.1161/01.res.0000027135.63141.89. PMID: 12142341.
22. Christoffels VM, Moorman AFM. Development of the cardiac conduction system: why are some regions of the heart more arrhythmogenic than others? Circ Arrhythm Electrophysiol. 2009 Apr;2(2):195-207. doi: 10.1161/CIRCEP.108.829341. PMID: 19808465.
23. Mommersteeg MTM, Dominguez JN, Wiese C, Norden J, de Gier-de Vries C, Burch JB, Kispert A, Brown NA, Moorman AFM, Christoffels VM. The sinus venosus progenitors separate and diversify from the first and second heart fields early in development. Cardiovasc Res. 2010;87(1):92-101.
24. Park DS, Fishman GI. The cardiac conduction system. Circulation. 2011 Mar 1;123(8):904-915. doi: 10.1161/CIRCULATIONAHA.110.942284. PMID: 21357845; PMCID: PMC3064561.
25. Rana MS, Christoffels VM, Moorman AFM. A molecular and genetic outline of cardiac morphogenesis. Acta Physiol (Oxf). 2013;207(4):588-615.
26. Yutzey KE, Robbins J. Principles of genetic murine models for cardiac disease. Circulation. 2007;115(6):792-799.
27. Hoffman JIE, Kaplan S. The incidence of congenital heart disease. J Am Coll Cardiol. 2002;39(12):1890-1900.
28. Bruneau BG. The developmental genetics of congenital heart disease. Nature. 2008 Feb 21;451(7181):943-948. doi: 10.1038/nature06801. PMID: 18288184.
29. Pierpont ME, Basson CT, Benson DW Jr, Gelb BD, Giglia TM, Goldmuntz E, McGee G, Sable CA, Srivastava D, Webb CL. Genetic basis for congenital heart defects: current knowledge. Circulation. 2007;115(23):3015-3038.
30. Goor DA, Lillehei CW. Congenital Malformations of the Heart: Embryology, Anatomy, and Operative Considerations. New York: Grune & Stratton; 1975.